
Dans l'article suivant, Jean-François Vibert décrit le système nerveux central en tant que machine de traitement de l'information. Cette architecture est illustrée par le cas, bien connu maintenant du système visuel, en particulier des premières couches de celui-ci (cortex visuel). Plusieurs niveaux d'organisation apparaissent, de la rétine aux cellules hyper-complexes. Toutefois cette simple hiérarchie ne suffit pas à rendre compte de l'extraordinaire efficacité du système visuel.
Jean-François Vibert
Le système nerveux central (SNC) est une formidable machine de traitement de l'information : extraction de signifiants, reconnaissance de formes, prise de décisions. De plus, il est peu sensible aux pannes et au bruit. Le système visuel est particulièrement représentatif sur ce plan, puisqu'il est capable de reconstituer une image mentale complète à partir de données parcellaires ou déformées.
Arriver à comprendre comment fonctionne le SNC pour obtenir de telles performances est un des défis fascinants de cette fin de siècle. Il intéresse les neurobiologistes, les théoriciens et les ingénieurs. L'actuelle explosion de travaux concernant les réseaux neuronaux en est une des manifestations.
Plusieurs niveaux d'organisation peuvent être décrits concernant les structures nerveuses, allant du moléculaire au niveau le plus élevé : l'intégration des données conduisant à la prise de conscience du monde extérieur, via de nombreux capteurs.
Nous essaierons de faire prendre conscience de l'importance de ce type d'organisation en niveaux, en prenant comme exemple le système visuel.
L'œil est un prolongement du cerveau, et la rétine est une partie du SNC, puisqu'elle dérive du neuro-ectoderme.
Une image est la distribution fugitive et instable d'une représentation 2D de l'espace extérieur.
Voir c'est déduire à partir de 2 cartes 2D, l'espace 3D qui permettra de se déplacer, saisir un objet, reconnaître des formes.
Le stimulus lumineux est le photon dont la quantité va donner la notion d'intensité lumineuse, et la longueur d'onde du rayonnement lumineux dont la valeur va donner la notion de couleur. Le spectre de lumière visible s'étend du violet (L= 400 nm) au rouge (L = 750 nm).
La rétine tapisse le fond de l'œil. Elle présente d'importantes différences régionales, ayant des conséquences fonctionnelles. Le centre de la rétine est dévolu à la vision colorée (photopique) et précise, alors que le pourtour est dévolu à la vision achromatique (scotopique) et plus globale.
On trouve 2 catégories de cellules nerveuses au niveau rétinien : les cellules de la voie directe (photorécepteurs cônes et bâtonnets cellules bipolaires et cellules ganglionnaires) et celles des voies indirectes (cellules horizontales et 3 types de cellules amacrines). Les cellules de Muller, et les cellules pigmentaires constituent le tissu de soutien et nourricier. On trouve de l'intérieur à l'extérieur, les cellules ganglionnaires, les cellules bipolaires, les photo-récepteurs et les cellules pigmentaires.
Il y a environ 160 millions de photorécepteurs par œil. Il existe 2 types de photorécepteurs : les bâtonnets dévolus à la vision achromatique (noire et blanche) ou scotopique (ce sont les plus nombreux) et les cônes dévolus à la vision chromatique (colorée) ou photopique. Il existe trois types de cônes, respectivement sensibles au bleu, vert et rouge.
Les photorécepteurs sont constitués d'un segment interne contenant le noyau et la terminaison synaptique, d'un segment intermédiaire, contenant un cil connecteur constitué de 9 paires de microtubules et d'un segment externe contenant le pigment photosensible.
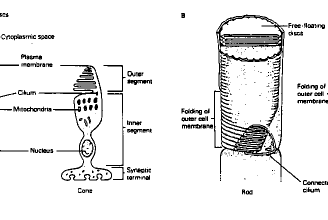
Schéma 1 Morphologie des bâtonnets et cônes
Le segment externe des bâtonnets a une forme allongée et est constitué de disques libres empilés, alors que celui des cônes est cônique (!) et constitué d'une membrane repliée sur elle-même pour former des invaginations horizontales (Fig. 1).
Les bâtonnets sont situés plutôt en périphérie rétinienne, alors que les cônes se trouvent au centre de la rétine. Le rapport nombre-de-bâtonnets/cellule-ganglionnaire est beaucoup plus élevé que le rapport nombre-de-cônes/cellule-ganglionnaire qui est voisin de 1. En effet, de nombreux bâtonnets convergent vers une même cellule bipolaire. tableau 1
La sensibilité des photorécepteurs à l'énergie photonique est due à la présence d'une substance chimique appelée pigment. Elle est localisée dans le segment externe. Le pigment des bâtonnets est la rhodopsine, celui des cônes est l'iodopsine. En fait, il existe 3 iodopsines, sensibles au bleu, vert et rouge, définissant 3 types de cônes selon la proportion de pigment de chaque type. En effet, les cônes contiennent les 3 pigments, mais par exemple, un cône bleu contiendra une molécule d'iodopsine rouge et verte pour 100.000 de bleue. C'est la proportion de molécules rouge/vert/bleu excitées qui compte, et 2 stimuli de compositions spectrales différentes paraissent identiques si ce rapport est conservé.
Le spectre d'absorption de la rhodopsine s'étend de 400 à 700 nm, avec un maximum à 490-500 nm (bleu-vert). Les bâtonnets sont 1000 fois plus sensibles que les cônes rouge et vert, ce qui explique leur rôle dans la vision nocturne (scotopique). Celui de l'iodopsine bleue s'étend de 400 à 500 nm (max : 470 nm). Celui de l'iodopsine verte s'étend de 450 à 650 nm (max :570 nm). Celui de l'iodopsine bleue s'étend de 550 à 750 nm (max : 600 nm). Les cônes bleus sont 5000 fois moins sensibles que les vert et rouge.
La transformation de l'énergie photonique en énergie électrique nerveuse sera étudiée en prenant la rhodopsine comme exemple.
Il y a environ 109 molécules de rhodopsine par bâtonnet. Elle est constituée d'opsine, protéine de 348 AA formant 7 hélices * transmembranaires emprisonnant un 11cis rétinal, forme aldéhyde de la vitamine A.
Un seul photon suffit à transformer le 11cis rétinal en 11trans rétinal en modifiant sa forme stérique. Par une chaîne de réactions (voir Fig 2), l'excitation du 11cis rétinal transforme les molécules de 3'5' Guanosine Mono Phosphate Cyclique ou GMPc en 5'GMP. La chaîne des réactions confère un pouvoir amplificateur important, puisque 1 photon produit 140.000.000 de 5'GMP.
Celui-ci agit en tant que second messager car il est annexé aux canaux sodiques de la membrane externe. Lorsque le cycle est fermé (GMPc) les canaux sodiques sont ouverts, et lorsque le cycle est ouvert (5'GMP), les canaux sodiques sont fermés.
Le fait que les canaux sodiques soient ouverts à l'obscurité (GMPc), fait que les photorécepteurs sont en permanence dépolarisés, et qu'il existe une boucle de courant sodique d'obscurité, avec un efflux de Na+ actif au niveau du segment intermédiaire et un influx de Na+ passif au niveau du segment externe.
Les récepteurs qui représentent le niveau cellulaire, présentent une réponse analogique, c'est à dire qu'ils répondent de façon proportionelle à intensité lumineuse (ils ne suivent pas la loi du tout ou rien).
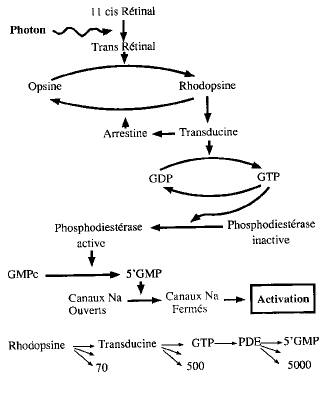
Figure 2 : la chaîne des réactions
La transmission vers les cellules bipolaires est probablement glutaminergique pour les 2 types de réponses, sans seuil, ni spike.
tableau 2
On distingue 2 types de synapses entre les photorécepteurs et les cellules bipolaires : celles conservant le signe de la réponse du photorécepteur (disfacilitation) et celle l'inversant (disinhibition). Les cellules bipolaires ne génèrent pas de spike (Fig. 3).
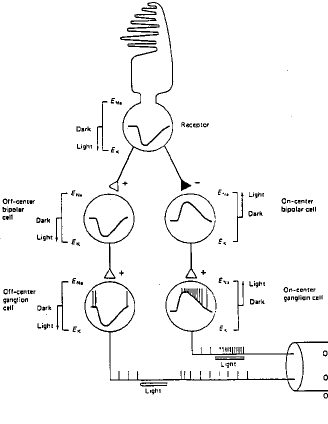
Figure 3 : les voies photoréceptrices et les réponses des différents types de cellules
On distingue 5 types fonctionnels de cellules bipolaires, un à champ large, recevant de plusieurs bâtonnets, et 4 types de cellules bipolaires pour les cônes (invaginantes et plates, à champ large et étroit).
Les cellules ganglionnaires génèrent des spikes. Elles peuvent être toniques ou phasiques. Chez le chat, les plus petits champs récepteurs font 120 µm de diamètre, correspondant a un rond de 1 cm de diamètre vu à 2 m.
Celles qui reçoivent des cellules bipolaires préservant le signe arrêtent de décharger à la lumière (off center).
Celles qui reçoivent des cellules bipolaires inversant le signe déchargent à la lumière (on center) (Fig.3 ).
Les cellules ganglionnaires on center déchargent lorsque leur champ est allumé, et cessent de décharger lorsque la périphérie de leur champ est éclairé (expérience de Kuffler 1952, fig.4 ). Les cellules off center présentent une réponse inverse. Ce comportement, dû à l'existence d'une inhibition latérale augmente le contraste.
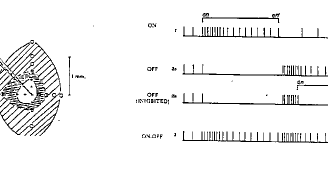
Figure 4 : l'expérience de Kuffler et les réponses des cellules ganglionnaires
Il existe 3 catégories de cellules ganglionnaires, correspondant à des fonctionnalités différentes. L'exposition prolongée à la lumière épuise les réserves de 11cis rétinal, et de transmetteur. L'adaptation photochimique (resynthèse de trans rétinal), se fait en 1h, alors que l'adaptation nerveuse (recaptage de glutamate) se fait en quelques minutes.
Les cellules horizontales font synapse avec les photorécepteurs. Elles modifient le champ de l'inhibition latérale et contribuent donc à l'antagonisme centre/périphérie. Le transmetteur est la dopamine. Elles n'entrent en action que si une grande surface est éclairée.
Les cellules amacrines sont situées dans la couche plexiforme interne, et constituent un niveau de traitement supplémentaire résumé par le tableau suivant.
tableau 3
En résumé, la rétine présente des caractéristiques inhabituelles. Les photorécepteurs sont normalement excités à l'obscurité (canaux Na+ ouverts), et le stimulus lumineux module l'hyperpolarisation.
La synapse récepteur/bipolaire n'a pas de seuil, elle libère son transmetteur en permanence. Cette libération est modulable par d'autres cellules. Un même transmetteur peut produire sur la même cellule des effets différents en fonction du récepteur postsynaptique. L'information est transmise sans spike. L'information est prétraitée grâce aux cellules amacrines et horizontales.
Il existe à ce premier niveau un prétraitement rétinien de l'information, avec :
des différences régionales
rétine périphérique
* Vision nocturne
* Détection des mouvements ;
rétine centrale
* Vision diurne
* Détails fins ;
des cellules ganglionnaires (centre-pourtour)
qui ignorent ce qui est uniforme ;
qui détectent les discontinuités
* spatiales (luminance)
* temporelles (mouvement) ;
un précodage de l'information
détection des figures ;
filtrage spatial
* simple chez le mammifère car le traitement de la complexité est déporté au niveau cortical
* complexe chez le batracien qui est peu corticalisé.
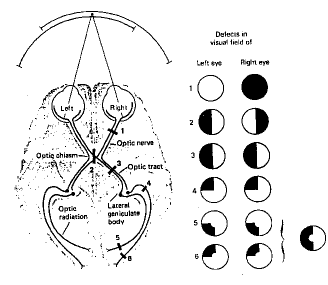
figure 5 : le croisement partiel des voies visuelles
Les informations qui arrivent de la rétine viennent faire relai au niveau des Corps Genouillés Latéraux ou CGL après avoir en partie croisé au niveau du chiasma optique. Les axones venant des hémi rétines temporales restent homolatéraux, alors que ceux venant des hémi rétines nasales rejoignent les CGL controlatéraux (Fig.5 ) et les Tubercules Quadrijumeaux Antérieurs (TQA) ou Colliculus Supérieur (CS). Il existe donc 2 voies visuelles qui vont emprunter des chemins différents et correspondre à des fonctionnalités différentes.
Au niveau des CGL, on a pu mettre en évidence, grâce à la technique du désoxyglucose marqué au C*14 l'existence de 6 couches, correspondant alternativement à l'œil droit et gauche.
Au niveau des CGL, les cellules présentent les mêmes caractéristiques fonctionnelles que les cellules ganglionnaires, avec conservation de la rétinotopie (représentation géographique de la rétine) et des informations fonctionnelles. On y retrouve les cellules X et Y, et la réponse centre/pourtour est conservée. Les axones des cellules des CGL rejoignent ensuite la couche 4c de l'aire visuelle primaire occipitale V1 ou aire 17. De là, les informations rejoignent les aires visuelles secondaires pré striées V2, V3 et V5, puis le cortex inféro-temporal. Cette voie constitue la voie de la vision focale.
Au niveau des CS les cellules se comportent comme les cellules ganglionnaires de type W, puisque ce sont les axones de celles-ci qui se projettent sur les CS. Les axones des cellules des CS rejoignent ensuite le pulvinar inférieur, un noyau du thalamus avant de rejoindre, soit également l'aire visuelle primaire occipitale V1 ou aire 17, soit directement les aires visuelles secondaires pré striées V2, V3 et V5. De là, les informations rejoignent le cortex pariétal. Cette voie constitue la voie de la vision ambiante (Fig.6).
Il est à noter que la rétinotopie est conservée jusqu'à l'aire corticale V1, avec une représentation très importante de la fovéa.
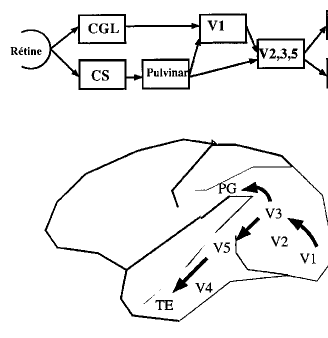
figure 6 : les deux voies visuelles
L'étude de l'organisation du cortex visuel a également bénéficié de la technique au désoxyglucose marqué. Celui-ci a permis la mise en évidence des colonnes de dominance oculaire. Sur chaque cortex, on observe une alternance de bandes de 400 µm représentant alternativement la rétine droite et la rétine gauche.
Au niveau du cortex, la réponse des cellules devient totalement différente de celle rencontrée jusqu'alors. Lorsque l'on descend une électrode perpendiculairement à la surface du cortex, on rencontre des cellules qui ne répondent qu'à une orientation donnée du stimulus lumineux (sauf au niveau de la couche 4c, où les cellules gardent le même type de réponse que les cellules du CGL). L'exploration systématique du cortex a mis en évidence l'existence de colonnes d'orientation d'environ 600 µm de large, organisées perpendiculairement aux colonnes de dominance. Dans ces tranches de 600 µm, on retrouve une succession de colonnes de cellules qui répondent toutes à la même orientation de 5 degrés en 5 degrés. Chaque tranche constitue une rotation complète de 180 degrés (Fig. 7).
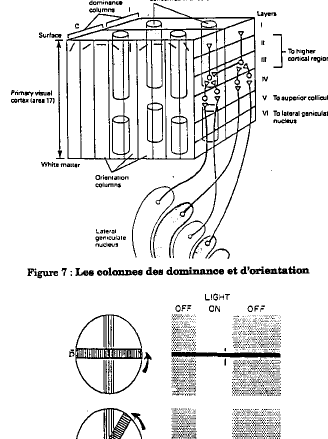
Figure 7 : les colonnes des dominance et orientation - Figure 8 : réponse d'une cellule simple
On appelle les cellules répondant à une orientation donnée, les cellules simples. Leur comportement résulte de la convergence des axones de cellules ganglionnaires physiquement disposées en ligne sur la rétine. L'orientation de cette disposition anatomique détermine l'orientation préférentielle de la cellule simple. Les cellules simples voient donc les lignes ou arêtes ayant une certaine orientation. On trouve là un nouveau niveau de traitement de l'information, par simple précâblage. On les trouve dans l'aire visuelle primaire V1 ou aire 17 (Fig.8 ).
Régulièrement réparties parmi ces colonnes d'orientation, on trouve des ilôts de cortex dans lesquels on ne trouve pas de cellules orientées, mais de cellules qui répondent à un stimulus coloré. On appelle ces îlots les blobs, ils font partie du canal de traitement de la vision colorée.
A la périphérie de l'aire 17 ou V1, on enregistre des cellules qui ne répondent que si un stimulus lumineux présente une orientation donnée et est en mouvement. La réponse est d'autant plus importante que le mouvement est rapide. On appelle ces cellules les cellules complexes. Elles résultent de la convergence sur une même cellule des axones de plusieurs cellules simples de même orientation et de champs visuels voisins. Elles utilisent le principe de la sommation temporelle : si on réexcite un soma peu de temps après une excitation précédente, la membrane est encore suffisamment dépolarisée pour pouvoir à nouveau atteindre son seuil avec un stimulus normalement infra-liminaire. Les cellules complexes voient donc les lignes ou arêtes en mouvement. On les trouve dans l'aire visuelle secondaire V2 ou aire 18. Elles constituent un nouveau niveau de traitement de l'information (Fig. 9).
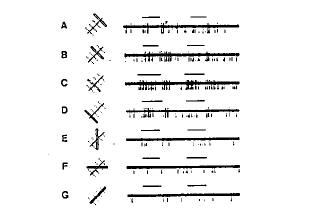
figure 9 : Réponse d'une cellule complexe
A la périphérie de l'aire 18, on enregistre des cellules qui ne répondent que si un stimulus lumineux présente un rapport surface éclairée/surface sombre donné, ou bien si un angle lumineux donné est présent. Là encore, la réponse est proportionnelle à la vitesse du stimulus. On appelle ces cellules, les cellules hypercomplexes. Elles résultent de la convergence sur une même cellule des axones de plusieurs cellules complexes d'orientations différentes et de champs visuels voisins. Les cellules hypercomplexes voient donc les formes en mouvement. On les trouve dans l'aire visuelle secondaire V5 ou MT ou aire 19. Elles constituent encore un nouveau niveau de traitement de l'information (fig. 10).
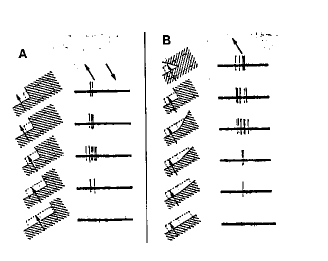
Figure 10 : Réponse d'une cellule hypercomplexe
Les aires 17 (V1), 18 (V2) et 19 (V5) contiennent les pièces de la perception. Elles obéissent au principe de la convergence croissante :
RETINE -> CGL ->SIMPLE ->COMPLEXE -> HYPERCOMPLEXE
A chaque niveau, chaque cellule voit plus que celle de niveau inférieur. Les cellules les plus hautes ont le pouvoir d'abstraction le plus élevé :
rétine et CGL -> position ;
Simples -> axe d'orientation ;
Complexes -> mouvement de l'axe ;
Hypercomplexes -> bords et angles.
Si l'on monte vers les zones de complexité plus grande (PG ou TE), on trouve même des cellules à très haute hiérarchie, capables de reconnaître un visage de face (et non de profil !).
On a vu que des portions spéciales du cortex, les blobs, sont dévolues à la vision des couleurs. On trouve, au fur et à mesure que l'on s'élève vers les régions à complexité croissante (aires 17 à 19), des cellules qui obéissent aux mêmes principes de convergence croissante. Mais les variables impliquées étant au nombre de trois (bleu, vert rouge), la combinatoire est également plus complexe. On trouve des cellules à opposition simple répondant au rouge au centre et au vert en périphérie par exemple ; des cellules à opposition double, qui sont inhibées par le vert au centre et le rouge en périphérie et excitées par le rouge au centre et le vert en périphérie, par exemple. On trouve des cellules répondant à une bande verte sur fond rouge, etc... Il existe également quelques cellules sensibles à l'orientation d'une ligne d'une couleur donnée. Ces cellules, sensibles à la couleur, constituent une voie de traitement parallèle à celle traitant la luminance.
Le traitement des informations par niveaux successifs de la rétine aux cellules hypercomplexes est générique, mais aussi subdivisé en plusieurs voies, qui fonctionnent en parallèle.
Elles partent de la rétine (parvo et magno cellulaires), et aboutissent au cortex visuel (V1 ou aire 17 et V2 ou aire 18). A ce niveau existent des colonnes (blobs) spécialisées dans la vision colorée.
Des aires V1 et V2 partent des connexions vers des aires intégratrices qui traitent la forme (V3), la couleur (V4) et le mouvement (V5) (Fig. 11).
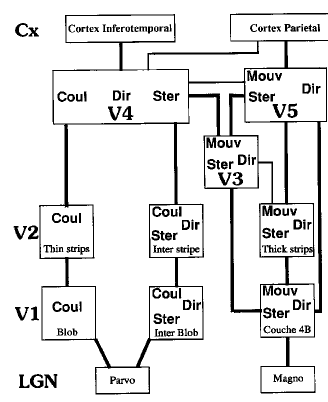
Figure 11 : les canaux de traitement parallèles
A partir des images sensorielles, le cerveau construit des images perceptuelles en combinant par convergence les notions élémentaires selon une stratégie schématisée par le tableau 4.
Finalement, comment les informations contenues dans toutes ces aires sont intégrées pour donner une vision unitaire du monde ? On ne sait pas bien ! Comme habituellement, il existe des mécanismes de rétro-contrôle importants : par exemple V5, spécialisé dans la détection des mouvements renvoit des informations vers V2 (cellules complexes), mais aussi dans les zones de V2 spécialisées dans la couleur et les formes (qui n'envoient rien à V5). De même V4 (couleur) se projette sur V2 (mouvements élémentaires). Il existe aussi des connexions entre les canaux parallèles : V4 avec V3 et V5, V5 avec V3 et V4, et l'ensemble reçoit des structures sous jacentes, venant de l'environnement, qui, par les propriétés de réseaux intègrent les informations non visuelles.
Cette intégration se fait in fine au niveau de 2 zones :
le cortex inféro temporal dévolu à la vision focale
. vision centrale
. reconnaissance des formes ;
le cortex pariétal dévolu à la vision ambiante
. vision périphérique
. position des objets
. mouvements.
Ainsi, la Nature a développé des structures et des stratégies permettant de passer du niveau du pixel (le stimulus lumineux ponctuel), jusqu'au niveau de l'image construite grâce à une convergence progressive des informations intégrées, en utilisant une économie de moyen remarquable, puisqu'il faut seulement 7 synapses pour passer du pixel à la forme !
Néanmoins, une simple hiérarchie de niveaux de traitements successifs ne suffit pas à rendre compte de l'extraordinaire efficacité de traitement de l'information effectué par le système visuel. Conjointement, plusieurs canaux de traitement fonctionnent en parallèle et convergent pour construire des images perceptuelles intégrant forme, couleur, mouvement, elles-mêmes intégrées à un niveau supérieur dans le contexte informationnel du moment, ou à l'activité en cours (adaptation des mouvements du corps à l'environnement, marche, préhension, etc...).
Bâtonnets Beaucoup de photopigment réponse lente grande amplification (1 seul photon) Réponse saturante non directionnel voies rétiniennes très convergentes grande sensibilité grande acuité achromatique : un pigment rhodopsine |
Cônes
peu de photopigment réponse rapide faible amplification réponse non saturante directionnel voies rétiniennes peu convergentes faible sensibilité faible acuité chromatique : trois pigments iodopsine (B, V, R) |
| type
W
X Y |
corps cellulaire petit
moyen gros |
arbre dendritique marge
petit large |
projection CS
CGL CGL |
rôle vision ambiante regard vision fine vision grossière |
| type
AI AII D
|
arbre dendritique large, très dense petit, dense très large, lâche
|
transmetteur Ach Ach Dopamine
| rôle
détection mouvement vision faible luminance module l'excitabilité générale |
| Image rétinienne
Intensité position x. y longueur d'onde temps œil (d/g)
|
Rôle sensoriel
Primaire luminance Spectre
Secondaire contraste vitesse 2D disparité Orientation 2D
|
Attributs inférés '1) Forme 3D forme taille rigidité
(2) Propriétés de surface couleur texture visuelle réflectance transparence
(3) Relations spatiales 3D positions relatives orientation 3D
(4) Mouvement 3D trajectoire rotation |
Dernière mise à jour : jeudi 23 octobre 2003
Dr Jean-Michel Thurin

|